During my undergraduate career, I had the opportunity to do undergraduate research with Dr. Katia Silvera at the University of California, Riverside. Dr. Silvera’s research focused on Crassulacean Acid Metabolism (CAM) photosynthesis of epiphytic orchid species. Epiphytic orchids have the flexibility to respond to environmental changes and utilize different photosynthetic pathways, ranging from C3 to facultative, to weakly expressed CAM and strong CAM. My contribution to this project was to prep samples of orchid leaves for stable carbon isotopic composition to assess the range of photosynthesis used by each species. I also compared the leaf anatomical patterns between orchid species with C3, weak CAM and strong CAM to differentiate the stomatal size and density among species. I found that CAM species had lower stomata density compared to C3 species in general and that CAM species had a correlation between the size of the stomate and the number of stomata per leaf area.
(a) Brassia gireoudiana with paracytic stomata, (b) Gongora leucochila with anomocytic stomata, (c) Kefersteinia costaricensis with anomocytic stomata,(d) Lockhartia micrantha with cyclocytic stomata, (e) Stelis argentata with actinocytic stomata, and (f) Trichopilia marginata with anomocytic stomata.

(a) Dendrobium kingianum with anomocytic stomata, (b) Guarianthe skinneri with anomocytic stomata, (c) Thrixspermum subulata with anomocytic stomata (d) Lophiaris silverarum with tetracytic stomata, (e) Vanilla planifola with anisocytic stomata, and (f) Rossioglossum ampliatum withanomocytic stomata.

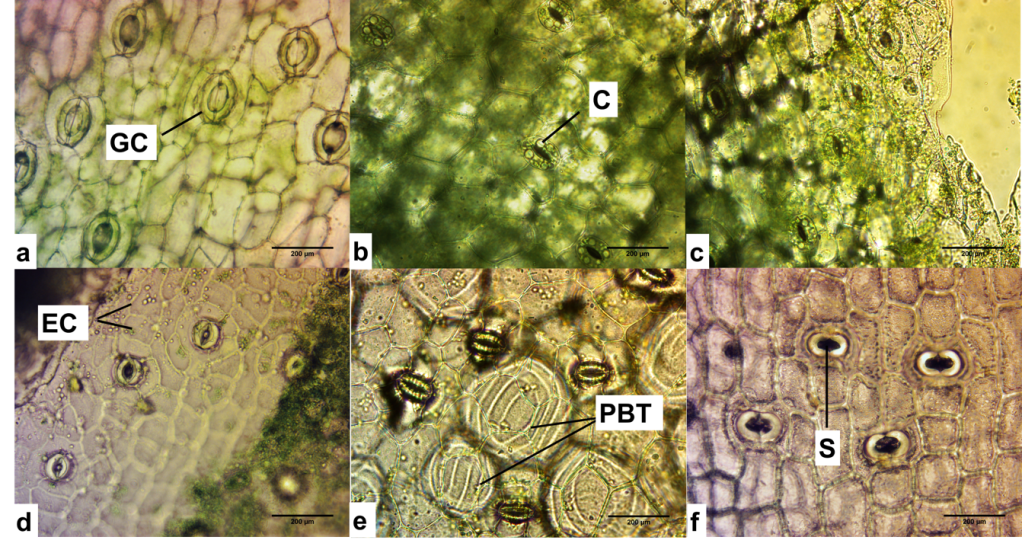
(a) Oncidium powellii with tetracytic stomata, (b) Oncidium sphacelatum with tetracytic stomata, and (c) Oncidium cariniferum with tetracytic stomata, epidermal cells (EC).
